| Apple Advanced Technology Group |
|
|
The second conference on artificial life (AL2) was held recently in Santa
Fe, New Mexico. The first AL conference (AL1) was held about two and a half
years before (Sept. 1987) at Los Alamos. Having attended both, it is interesting
to see the growth in both interest and active work in this essentially interdisciplinary
field. The first conference was attended by about 160 scientists and few
journalists; the second by about 340 scientists and quite a number of journalists.
For the most part, presentations at the first conference were deliberately
pedagogical in nature, in an attempt to communicate across disciplinary
lines; at this second conference many repeat presenters felt less need to
lay basic groundwork and jumped directly into the depths of their latest
research results. While this, perhaps, made some of the subjects a bit less
accessible to new attendees, it permitted the communication of a great deal
of new work, and may even have contributed to a sense of the emergence of
a true AL community.
To casual readers, I apologize in advance for the length of this document...
I went to this extent and level of detail in order to review the content
for my own research purposes, to fix it in my memory, and to serve as a
more durable form of memory for subsequent reference. If you wish an almost
vicarious experience of the complete conference, then I hope you read and
enjoy the entire document. If you just want to hit the highlights, you can
look for the "*" marks I have placed beside just the sections
that are either general overviews, especially interesting tidbits, or the
very best presentations. (In all instances, reading until the next bold
type will complete the marked section.) Note, however, that almost all of
the presentations were more than a little interesting.
* Artificial Life is defined by Christopher Langton (organizer of
AL1, co-organizer with Chuck Taylor of AL2) as "the study of synthetic
systems that exhibit behaviors characteristic of natural living systems.
It complements the traditional biological sciences concerned with analysis
of living organisms by attempting to synthesize life-like behaviors within
computers and other artificial media." Furthermore, it is "the
attempt to abstract the logical form of life from its material basis",
and "assumes life is more a function of the organizational form of
the matter than it is of the matter itself". All AL work rests upon
the fundamental assumptions that 1) it is possible to capture the essential
dynamics of living components, and that 2) simulated components utilizing
those dynamics will behave like real organisms.
Much of the work in AL is organized in a "bottom up" fashion,
focusing on local rather than global control, simple
rather than complex specifications, emergent rather than prespecified
behaviors, and, as with Braitenberg's approach in Vehicles, or Watson
and Crick's approach to determining the structure of DNA, concentrating
on modeling and synthesis as an alternative (and adjunct)
to the analysis of complex, frequently incomprehensible data. So
too was the conference organized in a bottom up fashion, with presentation
subjects moving from simulated physics, to chemistry, to origins of life,
to evolution, to development and learning, to ecologies, to societies and
cultures. The point was well made by Chris Langton, and by the work at this
conference in general, that such bottom up modeling not only has an intrinsic
value, but that acknowledging the various levels of organization and complexity
can help guide the selection of an appropriate level at which to model the
simpler rules that give rise to the complex, emergent behaviors observed
at higher levels. Just as simulating fluids at a "droplet" rather
than a molecular level is sufficient to produce many of the important and
interesting features of even a complex, turbulent flow, so modeling societies
at an individual level, or brains at a neuronal level may yield genuine
insights into fundamental living processes.
It is interesting that the Vivarium program has long had as its central
theme the idea of building ecologies in the computer, having under any
other name been engaged in Artificial Life research for a number of
years. While the purpose of the Vivarium program is frequently seen in terms
of exploring user interface and programming system concepts to support the
specification of agent or simulated creature behaviors, the method has been
and remains the development of simulated living organisms and ecologies.
Following are reviews - some brief, some fairly extensive - along with some
personal thoughts and comments, on the full set of oral presentations at
the conference, plus a few other conference events. The speakers are grouped
approximately according to the level of organization they addressed. A set
of abstracts, a few technical papers, an attendees list with addresses,
and my notes from the conference (plus the AL1 and Emergent Computation
(EC) conference reports referred to later) are also available through [no
one any longer] in the Los Angeles Vivarium office (xxx-xxx-xxxx).
Physics (a Meta-level applicable to all)
Tom Toffoli (MIT) - "Programmable Matter"
Described the beauty, or interestingness, of what we see as coming out of
the eternal battle between "exp vs. poly"; i.e., potentially exponential
growth in complexity of systems versus limited-resource-imposed polynomial
solutions and implementations. He suggests that systems exhibiting limited
communication neighborhoods and essentially identical function within each
of its components are the elegant solution achieved by nature in the exp
vs. poly battle.
He then described a series of computers designed at MIT specifically for
running cellular automata (CA's), called the CAM (CA Machine). The CAM implements
"programmable matter" where each cell may be thought of as representing
a single atom or particle. The dimensions and geometry of the cellular array
can be varied, as well as the connectivity and amount of state at each site.
He claims the existing CAM-6 represents about 1/30 of a Cray XMP, and describes
it as being a "scientific toy". The coming CAM-8 he claims will
equal as much as 1000 Cray XMP's, and still describes it as being a "scientific
toy", lamenting the overwhelming scaling problems in doing AL simulations
at the atomic level: a factor of 10^9 or 10^10 in going from molecules to
cells, 10^10 or greater in going from cells to organisms, and 10^6 or 10^7
in going from organisms to colonies.
* Chris Langton (Los Alamos) - "Life at the Edge of Chaos"
Chris gave a more mature version of the talk he gave at the Emergent Computation
conference (which I reviewed in an earlier report on that conference). Basically,
he has defined a lambda parameter, lambda, which is a ratio of the
number of generating states (the number of rules that cause a cell to turn
"on") in a CA's state transition table (the look-up table that
defines the CA's next state as a function of its current neighborhood) to
the total number of states in that transition table. This parameter, which
ranges from 0 to .5, encompasses the full range of behavior of a system
of CA's, from a system that immediately quenches at one extreme to a system
that is completely chaotic at the other. In between these extremes is a
regime characterized by local coherency and the transfer of information
between coherent patches, which he believes is the interesting behavioral
regime. These static and propagating structures provide the basis for embedded
computation.
He relates the range of behaviors he observes in his CA's to Wolfram's CA
classes: I - Homogeneous (fixed point), II - Heterogeneous (periodic), III
- Chaotic (non-periodic), and IV - Extended Transients (possible universal
computation), and to physical states of matter: I - Solid (globally ordered),
II - Liquid (locally, but not globally, ordered), and III - Gas (chaotic,
no order). By both entropic and mutual information measures made over a
large number of simulations he is able to demonstrate that there is a kind
of critical point around a l value of about 0.24, on the boundary between
solid and liquid, between type I and type II CA classes, where the type
IV CA class resides, where both persistent and propagating structures coexist,
that he calls the "edge of chaos".
It is only here, on the edge of chaos, that he believes living, information
processing systems have been able to evolve, and offers the interesting
conjecture that evolution is the process of learning to avoid attractors.
That is, systems starting out at too low a value of lambda always
fall into fixed, static, periodic attractor states; systems starting out
at too high a value of lambda always fall into chaotic, disordered
attractor states; but when the primordial porridge is just right, an extended
transient may be able to develop some local control over parameters affecting
its closeness to the critical transition value of lambda and so approach,
or at least stay close to that critical value, carry and process information,
and survive as a recognizable entity. Evolutionary steps that create new
organisms even more capable of tracking that critical lambda, and
thus avoid attractors, are successful and the organism becomes a better
information processor and survives more effectively, and so on.
* Chemistry and the Origins of Life
A significant number of presentations fell into this category. Most concerned
themselves with the origins of life, and with the formation, properties,
and dynamics of autocatalytic sets; that is, closed sets of molecules that
are able to chemically assist in the production of each other within a "primordial
soup" of constituent parts. Such autocatalytic systems are currently
felt to be the basis of primitive life, and the answer to the chicken/egg
problem posed by the DNA/RNA reproduction scheme employed by most life today:
for DNA to reproduce, complex proteins are needed, yet in living systems
these proteins are only synthesized by DNA - which came first? A current
belief is that primitive life began with an RNA-only scheme, and that RNA-based
autocatalytic sets provided a platform for the development of the more reliable
DNA/RNA scheme - a scenario which is lent additional credence by the recent
discovery that RNA can itself act as an enzyme/catalyst. Based on simulations,
graph theory, mean free energies, chemical reaction rates, and direct chemical
experimentation the various researchers attempted to define qualitative
and quantitative behaviors of complex chemical systems.
Steen Rasmussen (Los Alamos) - "Computational Chemistry"
Steen crafted an elaborate variant of the "Core Wars" game (utilizing
the same instruction set) that imposed energy/resource constraints on instruction
execution and used a couple of forms of mutation and natural selection to
evolve 'fitter' instruction sequences. He was never able to evolve self-sustaining
organisms from a raw chemical soup, but with a hand-tailored set of initial
conditions he was able to show waves of "chemical evolution",
in which different instructions tended to dominate the population. He likened
these behaviors to autocatalytic reactions in real chemical soups.
* Stuart Kaufman (Santa Fe Institute) - "Origins of Order: Self-Organization
and Selection in Evolution"
Stuart noted that traditional Darwinian views of natural selection tend
to ignore the process of self-organization. Stuart has done considerable
work (and presented formal analysis results at AL1) on the subject of autocatalytic
sets of polymer catalysts, and views the Origin of Life as an expected emergent
property of complex systems of polymer catalysts. That is, if you have a
sufficiently rich set of catalytic polymers, you will always find some set
of autocatalytic members; life is a natural, emergent property of closed
loops in randomly connected graphs. He sees spontaneous order and a "selection
to edge of chaos" (obviously agreeing with Langton) as the basis
of life. He also foreshadowed Danny Hillis's talk in stating that biological
evolution is coevolution, with evolving organisms forced to respond
to deforming, coupled energy landscapes, rather than simply optimize on
some static fitness/energy landscape.
He carried out a formal analysis of the expected number of attractors to
be found in a system of finite state automata (FSA) which use only canalyzing
Boolean functions (for which each input uniquely determines the function's
output state), such as govern nearly every genetic coding/biochemical reaction.
Suggesting that these attractors might correspond to viable cell types in
a living organism based on such functions, he determines that their number
scales as the square root of the number of FSA cells, and notes that the
number of cell types in living organisms also scales approximately as the
square root of the number of biological cells. The title of this talk is
also the title of Stuart's soon-to-be-published book, from Oxford University
Press.
J. W. Schopf (UCLA) - "What Must be True for Life to Have Evolved
on This Planet"
Basically a review of geologic time periods and the chemical composition
of the universe, the earth, and living organisms (noting the similarities).
One of the most interesting collections of observations was that oxygen
levels were < 1% of the current levels until after about 2 Ga to 1.8
Ga (Ga = Giga annum = billions of years ago); that anaerobic photosynthesis
not only does not require oxygen, but does not produce oxygen, while aerobic
photosynthesis both requires and produces oxygen; and that a variety of
complex chemical processes in the human body are anaerobic in early stages
and aerobic in later stages.
Vladimir Kuz'min (USSR Academy of Sciences) - "Symmetry Breaking as
a Possible Conducting Principle in Evolution: Example of a Chirality"
This was a fairly dense mathematical treatise on a particular form of symmetry
breaking (the usually unpredictable preponderance of one form of a seemingly
symmetric and equally likely pair of possible forms): chirality (or chemical/molecular
handedness). Language difficulties made it additionally difficult to follow.
However, I believe his main thesis was that symmetry breaking may often
be seen as the method of development from simple to complex systems. He
stated that life requires 1) the ability to self-replicate, and 2) handedness,
and furthermore, that 2) was required for 1). He claims that one of the
principle problems of biology is to explain the symmetry breaking of the
prebiotic world, noting that all proteins have one handedness, all sugars
the opposite handedness. He showed analytically (I think) that stable polymer
replication requires chiral purity (complete single handedness). And further
demonstrated that any simple "linear advantage factor" (in evolutionary
fitness, I believe) could not gradually break symmetry, so catastrophic
bifurcation is required. He also conjectured that such a linear advantage
factor would be essentially insignificant, and thus the sign of chirality
is purely accidental.
Ron Fox (Georgia Tech) - "Synthesis of Artificial Life"
Pointed out the inherent difficulties (the extreme magnitude) of digital
simulations of natural chemical systems. Also noted that "analog"
models are best based on actual molecules requiring the experimenter to
be a chemist. Observed that getting from the simple chemicals to the dynamic
processes of life is the crux of the matter, and suggests that an analysis
of energy flows may be adequate and appropriate to model evolutionary forces.
He pointed out that the interesting small molecular combinations
[H2O, Ca2+(aq.), H2PO4-, Na+(aq.)] have lower mean free energy than basic
elements, down to -3 DGf° (Kcal/gm). Quoting from his abstract: "Life's
origin may have involved a cellular structure capable of energy driven metabolism
at the level of coenzymes but without genetics or modern proteins. This
structure could provide the environment in which primitive genetics and
protein synthesis emerged based upon an RNA chemistry. This scenario leads
to the view that evolution has diversified polymer sequences over time,
rather than having selected a limited number out of a combinatoric plethora
present at the start".
* Doyne Farmer (Los Alamos) - "Protolife: Emergence and Evolution
of Robust Autocatalytic Sets"
Examining artificial chemical polymer sets using just ligation (condensation
or end-joining) and cleavage, Doyne carried out graph theoretic analyses
to predict the conditions under which autocatalysis will emerge. He was
able to determine critical transition conditions to autocatalysis as a function
of the radius (molecular length) of the food set, the probability of catalysis
(for any monomer pair to produce a polymer), and the number of "letters"
in his "chemical alphabet". Using assigned chemical reaction rates,
he was able to determine pumping rates (rate of introduction of "food"
chemicals - his environmental conditions) that resulted in a distribution
of most of the chemical mass into autocatalytic sets (as opposed to food
or background, non-autocatalytic molecular sets). He was then able to examine
the robustness of these autocatalytic sets by introducing perturbations
in pumping rate or alterations in the food set; some sets remained autocatalytic,
some didn't. He pointed out that these systems are lifelike in that 1) information
is passed on through time, 2) they grow more complex over time, and 3) they
depend upon diversity.
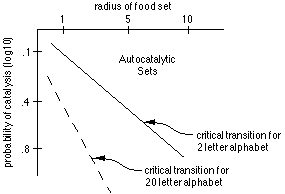
Stuart Kauffman made an interesting statement during the question and
answer session following Doyne's talk: if a completely general polymerase
emerges, then production of all nodes of the graph becomes possible; this
may be a critical threshold/development in the autocatalytic sets, which
essentially would give rise to the RNA/DNA structures.
* Peter Schuster (Universitaet Wien, Austria) - "Modeling Evolutionary
Optimization by RNA folding"
A really fascinating attempt to study "microscopic evolution"
by simulation of RNA folding. For their (with Walter Fontana now of Los
Alamos) study, the genotype consisted of strings of just 2 letters (as opposed
to the 4 letter sequences of real RNA/DNA), and the phenotype consisted
of the 2-dimensional folded molecular structure. The simple 1-dimensional
sequence of letters is referred to as the primary form, the folded 2-dimensional
shape as the secondary form, and a 3-dimensional folded shape (which their
study did not address) as a tertiary form. They modeled chemical strings
of length 70 with a 2 letter alphabet, represented by 0's and 1's, but using
the G-C interactions (I believe) to yield the chemical bonding energetics
that give rise to the secondary shape solutions.
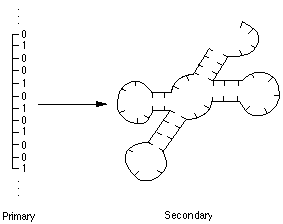
Populations of various genotypes are then evolved using crossover and
mutation, with fitness being determined from the phenotypical secondary
form (not from the genotype). Their evolutionary fitness function represents
a compromise between as many base pairs as possible and as small stacks
as possible (bonded pair sequence lengths, I think). From their simulated,
but fairly authentic chemical models they were able to determine that the
single strand elements (unbound loops) contribute to both higher replication
rates and to higher degradation (decomposition) rates. The "selective
value" (the phenotypic expression of the secondary form) varies largely
and abruptly with individual gene mutations, so the selective landscape
for these phenotypes is very ragged (with many local minima). There is a
critical mutation rate below which evolution is possible, with "quasi-species"
broadening as the mutation rate increases, and above which there is no evolution
- just random drift. The radical changes in phenotype with small changes
in genotype result in gradual changes in genotype, but radically different
secondary solutions and sharp changes in the phenotype during optimization.
Currently their crossovers may occur at any point in the primary string;
they believe that a more realistic model will emerge if crossover points
can be confined to the edges/ends of the already selected sequences.
This talk raised one of the most interesting issues for me, as I consider
the genetic code and embryology that will give rise to the "critters"
in my PolyWorld ecology-in-a-computer project. That is, contrary to conventional
wisdom, which says that small changes in genotype should result in small
changes in phenotype (supposedly in order to permit natural selection to
perform a graceful, gradient descent type of optimization), Schuster and
Fontana's work suggests that the norm may be for small changes in genotype
to produce radical changes in phenotype. This is also observed at a macroscopic
scale in nature, as evidenced by the Drosophila fruitfly, for which a single
gene's mutation can result in two pairs of wings instead of one (with all
the attendant reshuffling of the nervous system as well), possibly expressing
some genetic ancestor's now suppressed body structure.
Gerald Joyce (Scripps Clinic) - "RNA Evolution"
Though the work in Joyce's lab seemed to be aimed more at novel chemical
synthesis and novel synthesis techniques, their work clearly demonstrates
some of the fundamental principles of an "RNA world" - where RNA
is capable of serving as both genotype and phenotype, thus providing the
basis for an RNA organism. Using off the shelf chemicals they have used
tailored chemical reactions to provide a "selection pressure"
of 106 to 1 in favor of enzymes with a particular trait (a tail of a specific
molecular sequence), and in a closed cycle of selection, amplification (providing
suitable component chemicals and catalyzers to copy any and all enzymes
manyfold), and mutation, they have been able to engineer specific enzymes,
including a particular molecule (a mutant form of Tetrahymena ribozyme)
that cleaves DNA more efficiently than the non-mutated form cleaves RNA.
According to Joyce this engineered molecule is the first RNA enzyme that
has been shown to specifically cleave single-stranded DNA.
Ben Zuckerman (UCLA) - "Constraints on and Prospects for Life Elsewhere
in the Universe"
With a minimal definition of life (low entropy + complexity; which he pointed
out did not rule out automobiles), and a set of minimum conditions
to achieve such life:
1) flow of free energy
2) C+N compounds exist near the surface (or matter capable of interacting
with the free energy)
3) liquid H2O near surface (or a fluid medium for chemical transport)
4) no free O2 in the atmosphere (O2 being too reducing/chemically reactive)
5) time
Zuckerman delivered up a fairly pessimistic assessment of the possibility
of life elsewhere in our solar system (not much was said about life beyond
the solar system except that he sees little hope for life evolving except
on worlds similar to Earth). Venus fails due to too high pressures and temperatures;
Mars due to lack of water (the H2O channels we see are > about 109 years
old) and a highly oxidized, reactive surface; Jupiter due to high rate of
atmospheric convection (traversal of any hospitable vertical extent of the
atmosphere being so brief as to preclude the evolutionary time spans required
for even primitive organic chemistry to develop); Saturn for the same convection
problems. Uranus and Titan are more likely, though may not provide enough
free energy, and reaction times may be too slow. Io's surface is completely
recovered every 10 million years or so, yet is too cool for liquid sulphur
or silicates at the surface. He also speculated against the likelihood of
silicon-based life in general based on the (perhaps excess) stability of
silicon, its being less likely to make double-bonds than carbon, and the
fact that the Si-Si bond is only 1/2 the strength of the C-C bond. Sigh.
* Evolution, Development, Learning, and Ecologies
The steps on the ladder of organizational complexity do tend to blur a bit,
and most of the remaining presentations addressed some combination of limited
behavior modeling of individual organisms, evolutionary strategies for selecting
fitter populations of these individuals, and observing or attempting to
design or select for the emergent behaviors of groups of individuals. A
few papers addressed themselves to the ontogeny (individual physiological
growth/development) of plants and other organisms. There were also a couple
of talks and a panel on computer viruses, worms, etc. that actually sparked
quite a bit of discussion at the conference; in order to stick to a simple
chronological review of presentations these talks are lumped into this category
as well.
David Jefferson (UCLA) - "The Genesys System: Evolution as a Major
Theme in Artificial Life"
David (along with Chuck Taylor, conference co-organizer, and Robert Collins,
who later presented more recent results) characterized his approach as "organisms
as programs". The genotype of his creatures is a bit string, to which
GA operators are applied; the phenotype is the program - finite state automata
(FSA's) in one case, neural nets (NN's) in another. David attempted to evolve
creatures that were capable of exhibiting ant-like trail-following behaviors,
using FSA's in one series of experiments, NN's in the other. Genesys, their
simulator, is written in C++/Paris for the Connection Machine. For the FSA
critters, the genome is all bits of the state transition table; for the
NN critters, it is all bits of all weights. After each generation, they
reproduce a full population from the top 1% to 10% of the population in
a non-elitist fashion (not retaining the best organism, unless it
happened to not recombine and not mutate). They were generally using a 0.5%/bit
mutation rate, though he felt this might be a bit high. Their fitness function
was a numerical score based on how much of the trail the creature successfully
navigated within a fixed number of time steps (200, I believe), ranging
from 0 to 89. The 0th generation got a (maximum?) 58 score. With FSA's,
the 200th generation produced a full-scoring organism (a 14 state FSA).
With NN's, they got a full-scoring organism after 50 to 100 generations,
though the average score for both was only about 1/2 the maximum. Their
organisms clearly learned specific tasks relating to specific trails, and
when moved to a new trail learned worse (more slowly) than a fresh set of
random organisms (due to the fact that any small change in these pre-trained
organisms typically resulted in reduced performance and so was selected
against - this is a classic problem with trying to learn on an energy landscape
with a lot of local minima).
* Danny Hillis (Thinking Machines Corp.) - "Simulated Evolution
and the Red Queen Hypothesis"
The "Red Queen Hypothesis" of the talk's title is a reference
to the red chess queen in Alice Through the Looking Glass (Lewis
Carroll), who insists that Alice keep running, and when Alice says that
she doesn't see why she should, as she isn't actually getting anywhere,
the red queen replies that that is precisely the point... "it takes
all the running you can do to stay in the same place". (And it continues
something like, 'so to actually get anywhere you have to run faster and
faster and faster'.) As usual, Danny's talk was one of the highlights of
the conference. In support of the red queen's hypothesis, he showed and
discussed the results of some evolving systems he has been working with
on the Connection Machine. Genetic evolution as a simple optimization procedure
on a static, equilibrium fitness landscape, whether consisting of one or
many hills is really too simplistic a view. Dynamic, far from equilibrium
landscapes are the norm, perhaps necessarily so - as he demonstrates in
his "rampworld" and exchange-sort evolution experiments. Real
organisms must continually evolve to survive; it is as if every time some
species actually succeeds in becoming king of a particular hill, changes
in the environment (especially including the other organisms) cause the
landscape to shift, and the hill simply drops out from beneath the formerly
fit species. It takes all the running you can do to stay in the same place,
and you'd best evolve to run faster and faster and faster!
Danny used an example of attempting to evolve solutions to the creation
of crossword puzzles to discuss various problems with simple, single species
natural selection. Because of overlap in genes (words of different sections
of the puzzle overlapping in his crossword example), various partial solutions
are not compatible, so recombination doesn't help much, and organisms can't
get out of local minima. Mutation is (as is frequently the case) also inadequate.
Introducing "temperature" variations (fluctuations in the hospitability
of the environment) did not help - constant, "equatorial", benign
regions yield more diverse populations (as is seen in the real, biological
world). The solution that does work is to introduce a coevolving parasite
that specifically thrives on weak spots in the host population. As soon
as any variant of the host achieves a significant population size, by having
found some locally optimum solution to its fitness constraints, the parasites
begin to thrive and attack and reduce the host population, permitting alternative
solutions to emerge, build up a large population, and be attacked in their
turn...
Danny showed animated color contour maps of population densities evolving
over time for both the single-species and the dual-species simulations which
clearly illustrated the benefits of coevolution to both the average
and the best individual performance of the host species. These videotapes
were actually from a series of experiments that attempted to evolve the
best possible ramp (horizontal sequence of unit vertical steps); Danny also
talked about this experiment at the Emergent Computation conference, and
it is discussed in more detail in my report on that conference. He also
talked at that conference and this about evolving an optimum exchange sort
algorithm, a classic computer science problem; without parasitism the results
were unremarkable, but with parasitism he has been able to evolve the second
best (shortest) solution known to date.
An interesting observation about solutions evolved in this manner is that
in addition to providing a solution, it also provides a set of very adverse
tests for the solution (the best parasites), thus improving its reliability
and robustness. He noted that, "If airplanes flew based on sorting,
I'd rather fly on a plane based on this algorithm [the evolved one] rather
than one I drew up by hand".
In response to a question he also made an interesting, for my PolyWorld
considerations, almost offhand remark about the desirability of evolving
the embryology - a simple, but valuable idea to provide a more open-ended,
diverse system. He did however reiterate (from the EC conference) an opinion
that small changes in genotype should result in small changes in phenotype,
which seems to run counter to Peter Schuster's results and the drosophila
fruitfly example discussed above.
Alvy Ray Smith (Pixar) - "Simple, Non-Trivial Self-Reproducing Machines"
Alvy gave a "two page" formal mathematical proof of existence
for nontrivial self-reproducing machines, as cellular automata configurations,
which was based on the Recursion Theorem and relied only on computation
universality. He notes that earlier proofs (3 of them) have been book length,
very ad hoc, and depended upon "construction universality".
Jim Crutchfield (UC Berkeley) - "Evolutionary Mechanics: Towards
a Thermodynamics of Evolution"
Believes that "computational mechanics" (mathematics of nonlinear
processes, chaos theory) will provide a framework for modeling complex phenomena.
From his abstract: "The computation mechanics of evolutionary systems
suggests methods to quantify the temporal development of structural and
behavioral sophistication. The central thermodynamic variables, the statistical
complexity and dynamical entropy, measure the effective information processing
in an organism and in the environment. Variational principles govern their
change and suggest why and how complexity arises in natural systems."
He did not actually present any of these mathematical formulations at the
conference, but pointed us to a number of his technical papers.
Eugene Spafford (Purdue) - "Anatomy of Computer Viruses, Worms,
and Bacteria"
Spafford differentiated between worms (programs that can run on their own),
viruses (code segments that must insert themselves into other programs in
order to run and reproduce), and bacteria (or rabbits, which reproduce themselves
on the local machine - usually until some resource is exhausted - but do
not move from machine to machine). Most of Spafford's talk was a fairly
predictable elucidation of these invasive programs' taxonomy, structure,
and method of action. One frightening statistic that he quoted, I think,
was the existence of some 405 known viruses on PC's alone. And he claimed
that all of them are still active (noting that contaminated disks are frequently
filed away for years only to be finally hauled out to start a fresh round
of infection). He says that there is at least one example of a pair of viruses
that "mate" and reproduce offspring that are unlike either parent
(nVirA & nVirB).
Russell Brand (Lawrence Livermore) - "Computer Worms and the Turing
Test"
Russell gave an interesting talk (that heavily motivated audience participation)
based on a recent experience he had had in tracking down and identifying
a particular breach in machine security at the Lab. He listed a number of
the observed characteristics of the security breach, including an initial
large number of identical breaches, with no spelling errors, and simultaneous
activity on two different machines, which had them fairly convinced that
they were dealing with a worm. Additional observation yielded at least one
example of what appeared to be a typo, which was their first indication
that they might be dealing with a person or persons. For a considerable
period of time they were unable to determine whether they were dealing with
a worm or human(s), thus indicating the ease with which a program might
pass at least a 'no questions asked' Turing test. In the end, it turned
out that someone had written up this particular method of security breach,
made a number of photocopies, and passed it around; hence they were, in
fact, dealing with multiple people. He asked the pointed and interesting
question, "What is replicating here?" And answered it: not scripts,
not legends, not programs, not program fragments, but "memes"
(Richard Dawkins's word for replicating ideas).
Computer Virus Panel with Harold Thimbleby, Hyman Hartman, Eugene Spafford,
Russell Brand and David Jefferson
DJ asked if computer viruses are artificial life, and answered yes because
they are complex, self-replicating, demonstrate variation, mutation, recombination
and evolution, exhibit purposeful behavior, exist in a homeostatic (stable)
relation with the environment, tolerate perturbations in the environment,
have a metabolism (exchanging energy and/or information with the environment),
"live" in populations, and carry information.
* HT discussed his "Liveware" HyperCard stackware that
uses "friendly worms" to synchronize and maintain a database of
user interface researchers in Scotland. Liveware currently operates strictly
via 'sneakerNet', and functions by examining a newly inserted diskette,
then through its version control rules, incorporating data from any compatible
liveware stack on the new diskette into the host machine's database. (I
believe that the liveware stack on the floppy disk may be configured so
as to update itself as well.) Thimbleby stated that both liveware and worms/viruses
are autonomous and replicate, but where worms are uncooperative agents,
his liveware uses cooperative agents; that is, the behavior of his worms
is under the control of the user of the liveware. Actually, I think liveware
agents replicate data but not themselves and so may not actually qualify
as worms of any persuasion. In any event, I think his liveware work is extremely
significant and valuable, especially when extended to software agents that
may be used for synchronizing and maintaining the filestores on multiple
machines (he is currently working on a virus to keep several Mac filestores
in sync automatically).
Consistency is guaranteed in his liveware stacks by insisting that every
piece of data has a single owner, who is the only person authorized
to make changes to that data, thus providing a method for nonambiguous version
control. The current implementation of liveware definitely depends upon
friendly users, and has little if any security against willful misuse.
He provides users of liveware with a separate set of controls over the finding,
merging, and copying of data between stacks; each function may
be set to operate in one of three modes: manual, autotrigger, or
confirm. That is, upon insertion of a disk containing a compatible
liveware stack, liveware agents on the host machine may or may not automatically
carry out the data synchronization process, and may or may not ask for confirmation
prior to taking any automatic actions.
Thimbleby is currently looking for a publisher for his Liveware stack and
later developments as well. Jay Fenton of Farallon has expressed a possible
interest. Thimbleby has spoken to someone at Apple-UK, but doesn't feel
that is going to develop any further; if anyone from Apple-US reading this
has any suggestions or opinions about pursuing this further, you can (and
should!) contact him at (0786) 73171 or [email protected].
Even the current set of liveware tools seems interesting, and the filestore-maintenance
extension (especially with specific control over, say, folder-level synchronization;
carried out over networks and/or modems; and with some attention to security)
seems like one of the better ideas to come along in a while. (Wouldn't it
be nice if everyone could keep their own phone extension, mail-stop, department
information, and so on up to date in their own local copy of the Apple Phone
stack and have those changes propagate automatically to everybody else's
copy? Or if that special project folder and your expense report folder on
your home machine and your office machine could automatically stay in sync?)
At least Thimbleby appears to have implemented a reliable solution to the
version control problem (though I think that a finer granularity of ownership
and version control than he currently uses in his stacks might be useful
in a more general solution). I have some of his technical papers on the
subject if anyone is interested.
ES likened the seriousness of the crime of perpetrating a new computer
virus on the computer users community to that of someone introducing a new
biological virus that kills 5% of the human population of the planet. By
and large, of course, computer viruses are unquestionably a terrible thing,
but I think his analogy was more than a little stretched (and his paranoia
running more than a little rampant).
HH deliberately tried to offer a counterpoint to the rest of the
panel's theme, and discussed an interesting, still very speculative theory
that much of natural evolution's speciation may in fact have been mediated
by viral infection and an attendant alteration either in the infected organism's
DNA or in the process of copying it.
RB claimed that a virus is the wrong solution for any problem, and
supported his thesis by offering more efficient alternative solutions for
a number of possible applications of viruses.
An open discussion sought suggestions for defenses against viruses, and
yielded up the fairly obvious ideas of encrypted executables and anti-virus
viruses (against RB's better judgment, of course). Danny Hillis, however,
offered what I think was mostly a humorous speculation that we use different
operating systems on every machine; to make them practical we could evolve
them; and communication could proceed via a common interface. HH suggested
that the presence of viruses might force evolutionary growth of computers,
though I suspect that the only evolution one will see is to better protect
themselves against viruses, which will not necessarily be of any additional
value to anyone.
Tuesday Evening Demonstrations
Video and computer demonstrations were given of self-assembling models
of flagella rotors by Richard Thompson; a robot that bounces a tethered
ball by Brian Yamauchi; continuous growth graftal-like computer graphics
plants by Karl Sims, an AGAR-like (based on Minsky's Society of Mind)
behavior modeling/ecology in the computer system by Patti Maes; effects
of gain and neighborhood influence on logistic map functions by John Corliss;
microtubule self-assembly by Stuart Hameroff; neural net modeling of the
Vestibular Macula "accelerometer" in the head/visual system by
David Doshay.
Norm Packard (Santa Fe Institute) - "Complexity Increase in a Simple
Model for Evolution"
A fairly simple (deliberately he says, as he wants to develop an "Ising
model" for life) ecology in the computer. Rediscovered that crossover
is necessary to see significant evolutionary gains in fitness.
* John Holland (no current affiliation?) - "Echo: Explorations of
Evolution in a Miniature World"
This was a wonderful talk by the father of genetic algorithms, concerning
some totally new work (at least since the Emergent Computation conference
last May) he is carrying out entirely on a Mac II in his home. Afterwards
I asked him if he was being funded by Apple. He isn't. He should be! At
least to the extent of keeping him in the current fastest machine.
He describes this work as being "based on a gleam in Murray Gel-Mann's
eye". He has defined a grid world, with "resource fountains"
scattered around it, and inhabiting organisms who uptake resources from
the environment until they have enough and then they reproduce. On the face
of it, then, Echo is much like all the other ecologies in a computer. However,
Holland's resources come in a number of flavors (a,b,c), and are the same
string elements used to define the (variable length) genetic structure of
his organisms. Reproduction requires the accumulation of sufficient quantities
of all of the appropriate resources in an internal reservoir, so that the
genetic code can be copied letter for letter. Organisms random walk around
to get elements to reproduce.
Holland has also introduced a unique method of predation, based on dividing
up the genetic specification into "offense" genes and "defense"
genes (which can be and usually are different sequences of the resource
components/letters), and determines the outcome of organism confrontations
by cross comparing the two organisms' offense and defense "strategies"
(genetic strings). A score is calculated based on the number of matches
between each offense/defense pair, and the higher score yields a probability
of absorbing some resources of the loser. The loser's remaining elements
are returned to the environment. Each time step, each organism has a probabilistic
chance of entering into an interaction with a randomly selected organism.
Without this offense/defense split, selection is simply for the shortest
genome, using the most abundant elemental resources. With these organism
interactions, complex evolution emerges.
His next step is to make the genome serve as a grammar that generates marker
string tags (again, sequences of those same basic resource elements) for
the organisms, upon which interaction and preferential reproduction will
be based. When this is implemented he will have a genuine embryology (his
grammar) generating a true phenotype (the particular marker tags) from the
underlying genotype (element strings). He intends for the grammar to be
able to evolve like catalysts and classifier systems. With such a system
he hopes to be able to evolve special markings and recognition of those
markings in other organisms, mimicry, and other observable phenomena of
naturally evolved systems.
John ended his talk with some exhortations to researchers in this budding
field to: 1) seek answers to questions posed by target disciplines (don't
ignore the existing sciences); 2) try to use properties, interactions and
parameters relevant to the questions one's simulations are attempting to
address; and 3) use simulations in concert with appropriate mathematical
theories (to provide guidelines and predictions relevant to the model).
Finally he noted the need for a new "mathematics of perpetual novelty",
not equilibrium, for studying complex adaptive systems such as ecologies.
Robert Rosen (Dalhousi Univ., Nova Scotia) - "What Does It Take
to Make an Organism"
Attempted to relate some existing formalisms to AL. He also at times seemed
to be claiming that AL was a mathematical impossibility, yet then turned
around and offered "relational biology" as a new/old mathematical
formalism that can replace models based on the mathematics of traditional
physical sciences and have some hope of succeeding at modeling AL (I think).
Claims Rashevsky started "relational biology" (incorporating the
dynamical interactions with the environment in a total organism's model)
after a series of failures in biological modeling based on a reductionist
approach he dubbed "biomimesis". Rosen stated that "biology
is not simulable" because it is a "complex" system, meaning
that the fact that the "realization" (the "real" environment)
is directly influenced (in a Heisenberg sense, I believe he was suggesting)
by the "model" (our internal mental models of reality), as well
as the model being influenced by the realization, implies a lack of computability.
I think he's never heard of non-linear systems before. If you choose to
interpret his thesis as a semi-formal statement of the requirement for "grounding"
in the Cognitive Psychology sense it may have some merit.
Elliot Sober (affiliation?) - "Learning From Functionalism"
Sober was, in his words, the "token philosopher" at the conference.
He slipped back and forth between AI and AL, stating that "AI is to
psychology what AL is to biology". He noted that there is a distinction
between saying a computer model can help us understand life and saying a
computer model can be alive. He traced some historical developments
in the history of theories about the mind-body problem and claimed that
"Functionalism" can help yield insights in AL and AI (though I
never was exactly sure why). He discussed the Turing Test / Imitation Game
and noted the two interesting possible error conditions: Type 1 in which
the subject passes the test but in fact does not think (this is the most
common target for philosophical attack; e.g., Searle's Chinese story/Q&A
room and Block's tree of all reasonable conversations - the problem with
both these arguments being that they rest on the assumption of the actual
constructability of Searle's story-to-answers book and Block's conversation
tree). And Type 2 in which the subject fails the test but in fact is intelligent
(consider, for example, an artificial organism that is either simply uninterested
in mimicking human behavior, or is unable to pass due to a lack of shared
environmental and cultural experiences with humans but is still, in fact,
intelligent).
Sober simply read a prepared speech in a complete monotone, and basically
made everyone glad there weren't any other philosophers speaking (though
I remember how genuinely impressive I found the talk by Paul Churchland
- a Philosopher of Science - at the EC conference; see that report for more
info). He did offer up one good line with a bit of conviction (well, at
least vocal inflection), "If a system perceives, remembers, and responds,
then why ask if it can think?". But the rest of his talk was devoted
to asking anyway...
Ontology of Artificial Life Panel with Peter Cariani, Steen Rasmussen,
Norm Packard, Tom Toffoli, Robert Rosen and Elliot Sober
Generally one of the biggest bores at the conference, with everyone sitting
around trying to outdo the greatest philosophical thinkers of history by
deciding not only the nature and meaning of real life but the nature and
meaning of artificial life.
However, Steen Rasmussen gave a fun and elegantly succinct "proof"
of the possibility of AL and the benefits one might derive from it (shown
here in an only slightly edited form which Steen approved of - in fact he
kept my edited version of his handout):
(I) A universal computer is indeed universal and can emulate any process
(Turing).
(II) The essence of life is a process (Von Neumann).
Accepting (I) and (II) implies the possibility of life
in a computer.
(III) There exist criteria by which we are able to distinguish living from
non-living things.
(IV) If somebody manages to develop life in a computer environment which
satisfies (III) it follows from (II) that these life-forms are just as much
alive and you and I.
(V) If such an artificial organism perceives its reality, R2, for itself
R2 is just as real as our "real" reality, R1, is for us.
(VI) From (V) we conclude that R1 and R2 have the same ontological status;
although R2 in a material way is embedded in R1, R2 is distinct from R1.
(VII) Given (VI) it follows that it might be possible to learn something
about the fundamental properties of realities in general, and of R1 in particular,
by studying the details of different R2's. An example of such a property
is the physics of a reality.
* The other good bit from this panel was Tom Toffoli's description
of a Turing Machine (TM) with a second head. As he noted, the common, "individual"
point of view is to say that the first TM head (A) is basically a TM with
minor perturbations (due to head B's occasionally writing over head A's
tape marks). However, he points out, to really understand, to accurately
measure what is going on you must consider the complete, dynamical system
- which is not a TM. Correspondingly, a real, living organism and its environment
(all elements of that environment) comprise such a system.
Wednesday Evening Demonstrations
Marek Lugowski from Indiana University showed an interesting (though completely
ad hoc) tiling algorithm - some quite simple, completely local interaction
rules regarding which tiles can flip with other tiles - that permits a tiled
pattern to be completely randomized and then self-organize under repeated
applications of the algorithmic rules.
Mike Travers gave a nice demo of an upgraded AGAR in operation (his spreading-activation-amongst-mental-agents
behavior modeling system based heavily on Minsky's Society of Mind
and Tinbergen style Ethology). He demonstrated a more recent version of
his ant colony model that was shown at SIGGRAPH and figures prominently
in his Masters Thesis.
After another ecology simulator with Turing Machine-like critters called
"Turmites" (I missed the presenter's name), and a proposal for
another ecology simulator that was decidedly arcane and had never actually
been programmed up (by someone whose name I'm not displeased to have missed),
James Kalin gave a very nice and well received demonstration of SimCity,
the city administration game/simulator.
Peter Todd (Stanford) - "A General Framework for the Evolution of
Adaptive Simulated Creatures"
Peter and Geoffrey Miller are using a combination of NN's and GA's to provide
both learning and evolution in a simulated organism. They use bit-string
genomes that code for the type of connection between neurons, including
no connection and connections that are Back-Prop learnable, positive-only,
negative-only, or either, and, in recent versions of the simulator, unsupervised
(Hebbian learnable). Their system applies GA's in a fairly brute force way
to the entire network architecture, the connection strengths, types, and
so on. As one might expect, it learns, but slowly.
In their most recent experiments, the creatures roam around a grid world
looking for resources that are identifiable both by a "smell"
sense and a "vision" sense. Objects in the environment are equally
distributed between "food" (which increases the organism's available
energy 10 points and improves its fitness), and "poison" (which,
of course, has the opposite effects). Though in some of their simulations,
the organisms' vision system actually attempted to do pattern recognition
of 2x2 cell sized objects within a total 5x5 cell visual input pattern,
I believe they are simply attuned to select between one of two colors in
these experiments. The sense of smell is based on simple gradients of, again,
one of two odors. Food is always one color and poison the other during an
individual's lifetime, but the colors may change over evolutionary time
scales, and the creatures are provided with "100% accurate color transducers"
(their visual input neurons always are stimulated correctly according to
the actual color of the object). Smells, on the other hand, are different
for food and poison, and are fixed permanently (even over evolutionary time
scales), but the creatures' olfactory systems are given "noisy transducers"
(there is some probability that the olfactory input neurons may receive
reversed sensory input cues). The output from both the olfactory and the
visual sensory systems is combined to control the organisms actual response
to an object.
They deliberately tailored these sensory systems in the hopes that they
would evolve unsupervised, Hebbian learning (that is changeable over the
course of an individual's life, and learns directly from experience with
the environment) in the color/vision system, but essentially hard-wired
connections in the olfactory system that only change on evolutionary time
scales. They varied the accuracy of the smell transducer from 50% to 100%
accurate, and did indeed find a range between 70% and 90% smell accuracy
where their expectations for the emergence of different learning algorithms
were fulfilled, and "successful" organisms were evolved that combined
the two senses to give good survivability. With 100% smell accuracy they
evolved successful organisms, though as one might expect, evolution produced
a solution based entirely on smell. Interestingly, with 50% smell accuracy
evolution never produced a successful organism! Neither a useful smell-
nor vision-based solution could be found, and without the evolution of an
at least partially dependable "motivational/emotional/hedonic"
solution, an adaptive solution also was never able to evolve.
They made an additional interesting observation, based on earlier experiments,
that evolutionary pressure is first on the output motor effectors; motor
behavior was evolution's top priority (in order for species to accumulate
enough resources and live long enough to propagate).
The most interesting of their results to me was the failure to evolve successful
organisms at 50% smell accuracy. This suggests that advanced learning systems
may require an evolved innate ensemble of behaviors to use as a bootstrap
or platform for their own evolution. One of the PolyWorld goals is to explore
the possibility (desirability? necessity?) of using a hand-wired, innate
behavior based on an olfactory sense to guide the acquisition of learned
behaviors based on a vision sense [smell was left out of the version of
PolyWorld that actually got implemented].
Rob Collins (UCLA) - "ArtAnt: Evolution of Central-Place Foraging
Strategies in Ant-like Colonies"
Working with David Jefferson and Chuck Taylor, Rob reported some of their
most recent work based on evolving simulated ant colonies. They do their
simulations on a 1000x1000 grid world broken into 16x16 regions, with 435
units of food per region, 1 colony/region, and 4096 colonies, with 8 ants/colony.
The individual ants use pheromones (emitted by the environment, not the
ants if I heard him correctly?), food, compass heading, nest direction,
and a random number as inputs to a neural net with 1949 weights (note: real
ants have 100K to 200K neurons); the outputs are the allowable actions (such
as movement directions and food acquisition/disposition). I do not believe
that ants from one region were permitted to move into other regions. Each
generation was allowed to run for 500 time steps (about 25 min. on a CM2).
Reproduction is carried out at the colony level, not the individual
organism level, with the colony's chromosomes determined from Ant-0. The
colonies are scored based on food acquired less the amount of energy expended
on motion and on pheromones (so perhaps the ants do emit pheromones?). The
top 10% are selected and mated randomly. All "sister" ants of
the same colony are then different recombinations and mutations (at a 0.1%/bit
mutation rate) of the two colonies' chromosome sets.
Their ants did evolve to use their food sensors, though not optimally (they
frequently ignored close food). No ants ever evolved to use the pheromone
trails; they hypothesized that there was too much food available, hence
there was no selection pressure to evolve this capability. They want to
extend their experiments to more ants, with differing amounts of available
food.
David Stork (Stanford) - "Non-optimality via Preadaptation in Simple
Neural Systems"
David presented some results of evolving a simple simulated neural circuit
that models the crayfish tail-flip circuit. Particularly, he focused on
studying the effects of preadaptation of this neural circuit to perform
optimally for swimming and then adapting it based on performance of the
tail-flip behavior. Real crayfish do indeed use their tail both for swimming
and for this tail-flip, predator-avoidance behavior. Examination of the
neural circuitry in real crayfish provides a bit of a mystery regarding
certain connections whose function appears to be completely overridden by
alternate, inhibitory connections between the same neurons, or along the
same unique neural pathway. The reason for the existence of these useless
synapses is difficult to explain unless, David believes, one invokes a dependency
on historical function of the neural circuitry in question. That is, it
is well known that evolution and natural selection, by their nature, must
carry much of the existing baggage of an organism's physiology across generations,
making changes, sometimes subtle, sometimes a bit radical, but never so
radical as to make the new organism unfit to survive and propagate the changes.
Evolution was only presented with a Tabula Rasa once. David's hypothesis
is that a neural circuit that first evolved to swim optimally might very
well exhibit non-functional connections after being subsequently evolved
to tail-flip optimally.
David's neural models were fairly complex and biologically reasonable, based
upon the Hodgkin-Huxley equations. His genome consisted of the various parameters
to this model, and some method for expressing the neural network architecture
(number of neurons and connectivity). I am not clear about exactly how the
synaptic connection strengths were determined (particularly whether there
was any learning during an individual's lifetime - I think not), but he
mentioned that initial connection strengths were determined by similarities
in some of his genome parameters (presumably with previous generation circuits),
and reduced, initially, as a function of distance (from the pre-synaptic
neuron, presumably). The sign of a synapse (inhibitory or excitatory) was
determined by the particular pre-synaptic and post-synaptic neurotransmitters.
(He was also evolving the particular neurotransmitters being employed in
each neuron.)
His models were then trained for some number of generations with selection
pressure solely on the circuit's ability to produce a swimming motion in
the tail. These preadapted circuits were then run for a number of generations
with the organism's fitness changed to depend solely upon the circuit's
ability to produce the tail-flip motion. As he had hypothesized, the resulting
neural circuitry did indeed exhibit useless synapses, including some in
the same places as in the real network.
Someone during the Q&A session took issue with his particular technique
for evolving the neurotransmitters, especially a winner-take-all enforcement
of a single neurotransmitter per neuron (Dale's law - now definitely shown
to be incorrect), though I doubt he could do much else given the neuronal
model he was employing, or that this implementation detail in any way was
responsible for the primary results David observed. I do wonder about his
method of assigning connection strengths, and a lack of synaptic efficacy
variation in an individual's lifetime (though the latter concern may not
be an issue if this simple circuit is indeed non-learning in the real organism).
The largest concern I had with his model, that I voiced at the conference,
was that there was no selection pressure to reduce the circuit complexity
in his model (whereas biological systems do have at least some pressure
to reduce their metabolic energy expenditures). Indeed, his circuits typically
had a significant number of not only useless synapses, but useless - or
at least redundant - neurons. I think his basic thesis was entirely correct,
but his model was incomplete in a fairly crucial area.
Kristian Lindgren (NORDITA, Denmark) - "Evolution in a Population
of Mutating Strategies"
Kristian showed results from an evolving ecology of "strategies"
for solving the noisy, infinitely iterated Prisoner's Dilemma (PD) game.
That is, a single game consists of two opponents (the "prisoners")
deciding which of two possible actions to take ("Cooperate" or
"Defect"); the outcome of each of the four possible combinations
of decisions by the opponents is scored in such a way as to reward mutual
cooperation the most, single player defection (the other player must decide
to cooperate) second most, and mutual defection the least. In Kristian's
experiments, each PD game used a scoring strategy:
![]() player 2
player 2
![]() Cooperate
Cooperate ![]() Defect
Defect
![]() player
player ![]() Cooperate
Cooperate ![]() (3,3)
(3,3) ![]() (0,5)
(0,5)
![]() 1
1 ![]() Defect
Defect ![]() (5,0)
(5,0) ![]() (1,1)
(1,1)
The PD games are then repeatedly iterated, assuming a meeting between the
same two opponents (in which case a good strategy is "Tit-For-Tat";
that is, choose the same action as the opponent did last time). And introducing
noise means causing the actual performed action of a player to be opposite
to the intended action with some probability p.
Kristian then defines a variable length genetic code that determines a player's
decision based on some amount of history of both players' decisions. These
genetic codes are 0/1 (Defect/Cooperate) bit strings that code for a unique
decision for all possible histories (up to the depth permitted by the length
of the genome). The position in the bit string used to decide a particular
next play is determined directly from the numerical value of the historical
sequence of 0's and 1's (base 2, of course), and the value of the
bit in that position is the action to be taken. For example, to support
a history with a depth of 3 steps, 8 bits (2m, where m = historical depth)
are required in the genome, and a play sequence where the player in question's
opponent had most recently defected (0), prior to which the player
had cooperated (1), prior to which the opponent had also cooperated
(1) would produce bit position (110)2, or 6 in base 10, so the value of
the 6th bit in the player's genome would decide his next intended
action.
Point mutations and gene duplication rates of 2x10-5 and 10-5, respectively,
were used. Kristian noted that even though gene duplication doubles the
length of the genome, it does not change the phenotype (strategy); I think
this implies that the same number of historical steps still goes into each
decision and the longer genome is processed in chunks, with the player simply
having the ability to alternate decision strategies, as opposed to using
the greater historical depth directly, but am not sure. I do not recall
him mentioning crossover and recombination between players, so I suspect
that this simple system may have relied on mutation alone for improvement.
As Kristian showed with plots of population fractions, even this simple
system yielded some very interesting population dynamics. Starting with
just 2 bit (single step history) genomes, the fraction of the total population
occupied by the four possible phenotypes trades off in a complicated manner
for around 1000 generations, whereupon a stable coexistence dominated principally
by a Tit-For-Tat strategy (at about 60%) and a negative Tit-For-Tat strategy
(at about 40%) emerged. This remained in approximate equilibrium until around
5000 steps at which point complex dynamics recurred until around 12,000
steps a stable population of (0001) strategies with very low (00010001)
strategies emerged. This subsequently became chaotic again until another
stable configuration emerged at around 30,000 steps, and so on, with no
permanently stable equilibrium condition appearing as long as he ran the
simulations.
His point was primarily that this simple system can serve as an effective
testbed for studying some aspects of complex system dynamics, as evidenced
by the various emergent properties it exhibited, including the intrinsic
changes of dimensionality represented by the evolution of longer genomes
that were more successful than lower dimensionality players, the periods
of stasis with coexistence/mutualism, and population dynamics best represented
by a punctuated equilibrium.
Alvy Ray Smith (Pixar) - L-systems, etc.
Due to the absence of one of the intended speakers (Przemyslaw Prusinkiewicz),
Alvy held forth in an impromptu talk on Lindenmayer grammars (L-systems),
and their application to growing computer graphics plants. Some pretty pictures.
* Martin de Boer (Univ. of Utrecht, The Netherlands) - "Modeling
and Simulation of the Development of Cellular Structures"
Martin showed some interesting work, carried out with Przemyslaw Prusinkiewicz
and David Fracchia, that coupled some simple cell physics with two dimensional,
continuous L-systems to produce some strikingly realistic ontogenic development
patterns for algae growths, snail embryos, and ferns. They chose to model
2D growths for computational tractability and because the natural forms
could be directly observed (rather than killed and sectioned).
Martin referred to work by Lloyd which showed that specific sites of cell
division and attachment are inherited from the parent's microtubules ("inheritance
by cytoplasm" ! ). Martin et al developed L-system models, based on
observed and measured characteristics of real organisms, that included the
generation of special cell wall markers in the grammar. These markers were
used to determine cell division sites. Divided cell pairs directly replaced
the parent cell as their model 'aged'.
Simple cell physics were determined from an internal, osmotic pressure that
was calculated proportional to cell wall length, plus simple Hooke's law
springs for the cell walls. After cell division, these forces are out of
balance, so the system of cellular forces is allowed to relax to a new equilibrium,
then the cells divide again, and so on.
Granted the L-systems were tailored specifically to produce cell division
sites that correspond to the actual organism being modeled, but the resulting
simulated cellular development patterns were rather astoundingly like the
growth patterns in the actual organisms, including large, global features
such as overall external shape (matching a heart shape growth accurately
in one case). Though this method is a bit of a CPU-hog, it is the most interesting
simulated embryology/ontogeny that I have seen, and might end up playing
a role in PolyWorld.
Narendra Goel (State Univ. of New York) - "From Artificial Life
and Real Life - Computer Simulation of Plant Growth"
Goel is attempting to use context-free L-systems and computer graphics to
produce realistic models of plant growth. The work is currently 2D (necessarily,
as it is being done on PC's). They intend to correlate parameters in their
L-system specification to environmental factors, and thus be able to model
and illustrate the impact of these various factors on crop yield. They currently
do this by a completely proscribed sequence of changes to the L-system parameters
corresponding to a particular set of environmental conditions, but they
hope to develop an algorithmic model for relating environment to L-system
parameters. They also intend to specify computer graphic lighting parameters
that are carefully calibrated against actual spectroscopic light reflection
measurements made using real plants (though this aspect of the work is not
very far along as yet). He showed a surprisingly realistic looking corn
plant modeled with his system, and demonstrated the effects of different
environmental conditions on its growth. Everything is currently so hand-tailored
to produce this realism that it is difficult to assess how well their system
will be able to satisfy their longer term goals, but the images they showed
were quite impressive.
The Unknown Researcher (?) - "?"
Unfortunately I missed the presenter's name and the title of this unscheduled
talk... but he demonstrated the ability of "Iterated Function Systems"
(IFS, a finite set of "contraction mappings"; i.e., functions
for which it holds that | f(x2) - f(x1) | < | x2 - x1 |, for all x) to
produce images akin to L-system ferns, Sierpinski triangles, and other fractal
geometries. He also developed a formal proof of the ability of L-systems
to produce equivalent forms to IFS's.
Thomas Ray (Univ. of Delaware) - "Tierra: An Artificial Life Simulator"
Tom demonstrated an AL simulator that successfully evolved assembly language
instruction sequences. He gave each organism a set amount of CPU time in
which to execute its instructions, which effectively corresponded to the
organism's energy supply. He thought of the organisms as "cells",
whose size was a function of their memory allocation block size. He referred
to them as "semi-permeable" because a cell could not write into
another cell's interior, but it could read and execute another cell's code.
His assembly language did not support numeric operands, just registers,
though he made a comment about cells being able to "create a numerical
value in the CX register". He did not have time to effectively present
any details about his instruction set. He did show the system running in
real time, and noted the emergence of some of the classic ecological dynamics,
including mutualism and punctuated equilibrium.
Bruce MacLennan (Univ. of Tenn.) - "Synthetic Ethology as an Approach
to the Study of Language"
MacLennan listed the principle language issues as: How do languages emerge?
What are the supporting mental states? What constitutes intentionality?
How are worlds constituted by language communities? How can syntax emerge
from pragmatics? He then sought some answers from Wittgenstein: "Language
gains meaning from usage", Heidegger: "Concerns, expectations,
and practices of the community give language meaning", and Popper:
"Animal intelligence is continuous with human intelligence". I
believe he was also quoting Popper when he said, "The main task of
the theory of human knowledge is to see it as continuous with animal knowledge,
and the discontinuity - if any - from animal knowledge."
MacLennan believes we must relate the ethological context to the physiological
methods of communication. Behaviorist and ethological approaches both have
shortcomings, but by combining the simplicity and control of behaviorist
methods with the ecological and pragmatic validity of natural ethology he
believes we can develop a "Synthetic Ethology" that may serve
well in developing an understanding of communication and language. He suggests
making "REAL" worlds inside the computer; that is, don't think
of the computer as an abstract symbol manipulator, but as a mass-energy
manipulator - creatures in such a world are real. To define communication
he quoted Burghardt (I think), "Communication is the phenomenon of
one organism producing a signal that, when responded to by another organism,
confers some advantage on the signaler's group".
MacLennan then described a simple world he has simulated in which a number
of "simorgs" (simulated organisms) all have completely private
local environments which can only be read (not written), and read only by
the owning simorg. In addition, there is a shared, global environment which
may be both read and written into by every simorg. Thus any information
about another simorg's local environment can only happen through communication;
that is, by one simorg "emitting a sound" by placing a symbol
into the global environment that is received by another simorg. The simorgs
internal state is represented by a finite state automata. I was not able
to determine much about the actual nature of the local environments, nor
therefore the nature of the advantage conferred by communicating with other
simorgs about them, nor the complexity of the simorgs defining FSA's, nor
how learning occurred in this context.
MacLennan claims that genuine communication emerged between simorgs. He
next wants to see if he can understand the content and syntax of these communications.
One way in which he analyzes and documents the learning is by studying the
statistical characteristics of a "Denotation" matrix. This D matrix
builds a table where rows correspond to all possible states of the "local
situation", and the columns correspond to the "global communication"
(How? Is this the set of symbols? The set of all possible sets of symbols
containable by the global environment? Something else?). During the course
of the simulation he increments the appropriate entry in the D matrix whenever
a successful communication occurs (How is "successful" defined?
Is it simply that one simorg reads what another has written?). He states
that without learning the D matrix is fairly uniform and has high
entropy, and that with learning the D matrix is sparser and has lower
entropy.
Sigh. I really think there are some interesting aspects to the way
this experiment was designed, but as all of the questions above indicate,
its significance is difficult to assess due to an incomplete communication
between the speaker and this organism.
Rob J. De Boer (Los Alamos) - "The Development of an Immune System"
Rob comments that the immune system is a complex information processing
system embedded in a universe of protein patterns. He notes its ability
to carry out recognition tasks in its specificity for particular antigens,
its use of long term memory mechanisms to grant immunity that lasts for
months and years, its use of evolutionary mutation and selection in antibody
maturation to adapt its response to antigens, and its ability to classify
self versus foreign that emerges from a selection process during neonatal
life. He and Alan Perelson have carried out simulations that explore the
selection process that in human beings results in a stable population of
some 106 different types of lymphocytes even though more than 1012 different
types are producible by the immune system. Their simulation results agree
well, at least qualitatively, with the real immune system's behavior.
Pauline Hogeweg (Univ. of Utrect, The Netherlands) - "On the Natural
History of Artificial Life"
Pauline exhorts ALife researchers to examine and learn from the Natural
History of their simulations so that they may glean some of the same insights
that the study of Natural History provides for natural biotic life (hereinafter
dubbed "BLife"). She discussed an extremely simple rule set for
modeling chimp behavior - that basically will just generate behaviors to
seek out either food, or a mate, or a group at every time step - and claimed
that a simulated ecology full of these simple chimp models will produce
population statistics very like ethological data on chimps. She suggests
that the observed ethological behaviors emerge in direct response to simple
environmental parameters. (Of course, the parameters in her model were selected
specifically to produce the behaviors they agree so well with, so who knows?)
Rik Belew (UCSD) - "Evolving Networks: Using the Genetic Algorithm
to Design Connectionist Networks" changed to "Models of
Learning and Evolution"
Rik discussed, among some other observations on the ALife research field
in general, some of the work he has been doing in using GA's to evolve at
least some optimal learning rate parameters for Back-Prop (BP) NN's. The
GA did indeed come up with a set of parameters (learning rate of 3.0 and
momentum term of 0.3) that seem counterintuitive and yet dramatically improved
the NN's learning rate, at least for the problem he was attempting to solve.
He also indicated that the GA+BP combination succeeded in producing dramatically
more accurate network solutions than either the GA or BP on their own (which
suggests he has also been using his GA's to adjust the network weights rather
than just its learning rate parameters). This is principally due to the
combining of the GA's principle strength of effective wide sampling (on
a much more efficient basis than simple random sampling) with BP's principle
strength of good local optimization.
Stephanie Forrest (Los Alamos) - "Using GA's to Study the Evolution
of Cooperative Behavior"
Stephanie discussed some work she has been engaged in using GA's to try
to evolve arms spending strategies that will result in a stable balance
of power amongst three countries. A rule that "the two weakest countries
are always allied" is enforced, and the new year's arms spending is
a fairly simple algebraic function of last year's spending, an "intrinsic
self-armament level", and some rate constants. Their fitness is determined
by the absolute magnitude of the difference between the expenditures by
the strongest country less the sum of the two weaker countries expenditures.
(Stephanie notes that later they would like to also minimize the total expenditures.)
The results to date show a regular basin of attraction (a stable arms spending
strategy) when fitness is precisely at 0 (optimum for the way their fitness
is defined), however, with the slightest deviation from this optimum (a
fitness of 0.002 is sufficient), the various countries' arms spending policies
fluctuate radically.
She has a lot of good ideas for improving the model in the future, including
evolving the countries' policies on the same time scale as global interactions,
representing each country by its own population of (selfish gene) bitstrings,
each country deciding independently how to allocate resources and form alliances,
and making fitness dependent on internal national stability and interactions
with the other countries.
* David Ackley (Bellcore) - "Learning from Natural Selection in
an Artificial Environment"
Thanks to both some genuine technical innovation, and a sense of humor,
this was truly one of the high points of the conference. As David (and Michael
Littman) point out in their abstract: "The process of natural selection
is clearly a source of information about the performance of an individual
organism, but -- since the signal for failure is death -- it is not immediately
apparent how it could be exploited to perform learning during an individual's
lifetime." Their clever solution to this enigma, which they call Evolutionary
Reinforcement Learning, was to evolve a moment-to-moment evaluation function
within the organism that is subsequently able to provide a moment-to-moment
reinforcement signal. They used separate NN's for the organism's behavior
model and for its moment-to-moment evaluation function. Then, in order to
be able to effectively apply this rapid reinforcement signal and to take
advantage of a reasonably well understood NN learning algorithm (Back-Prop),
they invented another clever technique that they call Complimentary Reinforcement
BackProp (CRBP). CRBP works by using the reinforcement signal and the previous
activation levels in the output layer of the network (in this case the behavior
modeling network) as a probabilistic generator of a training signal for
that output layer (resulting in a vector of desired activation levels which
are all either at full activation or no activation). Simple BackProp is
then used to update the connection strengths in the behavior modeling network.
So, the rapid evaluation function is evolved over multiple generations under
selection pressure provided by the life-or-death/once-per-generation evaluation
function, and the behavior model learns as best it can from whatever the
current version of the rapid evaluation function is able to tell it.
David presented a wonderfully informative and funny videotape that
describes their artificial life environment, called AL. Again from their
abstract, their computer simulations "span four orders of magnitude
in space and six orders of magnitude in time. Successful individuals may
achieve lifetimes of 25,000 steps or more, and initial populations that
develop long-term viability may descend through 300 generations or more
before arriving at the one million step simulation limit."
I will attempt to obtain a copy of this videotape for public consumption;
it's a delightful examination of some excellent work.
John Nagel (affiliation?) - "Animation, Artificial Life, and Artificial
Intelligence from the Bottom"
John noted that AI has not succeeded very well with the top-down approach.
And had lots of reasons to be encouraged by the AL approach. Quoted from
or referred to lots of people's work, including Michael Kass's Luxo lamp
animations, Mike Travers's thesis, and Terry Winograd ("The hard thing
is deciding what to do in the next 15 seconds."). Recognizes that human
level AI is too lofty a short term goal, that even a squirrel might still
be too hard, and proposes a squirrel that does the right thing over periods
of less than a minute. Quoted Hans Moravec's estimate of 1 gm of brain mass
being equivalent to about 1000 MIPS (humans would then be 106 MIPS), but
was taken quite severely to task for this by Maureen Gremillion of Los Alamos
(who vehemently proclaimed that Moravec's theories are completely wrong
- based on her experience in computer modeling of the human visual cortex).
Really this entire talk was pretty content free.
Rod Brooks (MIT) - "Real Artificial Life"
Rod showed some fun robots that he and his lab have been developing over
the years. All were based on Finite State Machines. All behaviors were built
using a layering technique wherein each layer represents a single, simple
behavior, and modifies lower level layers through a relatively simple inhibition/excitation
mechanism just prior to passing on the signals to the motor actuators. For
example, level 0 is usually avoid collisions, level 1 may be to seek motion,
and so on. Few if any of the robots had more than 3 seconds of memory. Among
the numerous robots he discussed and showed video clips of were a robot
that sought out, picked up, and returned empty coke cans (called, of course,
the "Collection Machine"), and a robot that sold candy and then
used its profits to bribe people to open doors for it (called, yes, the
"Confection Machine"). They have experimented with 6-legged insect-like
walking robots that are able to traverse rough terrains of jumbled textbooks
and so should easily survive the less educated surface of the moon. Rod
noted the progression of robot masses over the last few years, from the
Confection Machine which weighed in at 50 Kg, to a machine called Squirt
that weighs just 50g, to some work in progress now on a 1mm diameter robot
with 20 legs. Rod speculated that robots of approximately this size might
be able to be constructed that fed off of electrons, lived in the corners
of TV screens, and would wipe the dust off of the screen when the set was
turned off.
Alan Kay (Apple) - Vivarium overview
Since the Vivarium program, as pointed out earlier, has always had an essentially
AL central theme, Alan gave a very appropriate overview of the history of
the Vivarium program. He pointed out the original concept started with Ann
Marion's simulated aquarium, and noted that our goal is still to provide
children with something like a simulated aquarium where they may create
both the form and the behaviors of their animals, place them into the "aquarium",
and see things from their own animal's point of view.
In terms of the animals' forms, the intention has been to take advantage
of the conformally related body shapes shared by many fish. To approach
the animals' behavior modeling Alan referred to Grey Walter's The Living
Brain and talked about the "turtles" Grey built, Elmer &
Elsie. He noted that as their thinking progressed, he and Ann had to begin
considering methods for modeling emotions (briefly discussing the Lorentz
fluid theory of emotional discharge), and also change their thinking from
a simple "kit" to a very general environment builder.
The work to this point had been going on at Atari, but it was at this point
that Atari essentially folded. Alan talked briefly about some of the explorations
at the MIT Media Lab, and then jumped to two years later when they discovered
the Open School in Los Angeles. He described the school (300 children, 10
classrooms, 60 children per cluster, 2 teachers per cluster, clusters are
2 grade levels), and noted that there are about 200 Mac's in the school,
which are in use about 2 to 2 1/2 hours per day. After a brief flirtation
with VideoWorks, HyperCard was released and rapidly became the lingua
franca (sic) for the entire school.
A second strand running through the current incarnation of the Vivarium
program is PlayGround, the language and interface that we hope will permit
children to define agent/animal behaviors in a simple, straightforward fashion.
Alan noted that PlayGround had been under development for a couple of years
by Jay Fenton, himself, and Ann Marion, and mentioned testing it in the
Open School.
Alan briefly talked about some of the other projects within Vivarium, including
the machine learning work by Larry Yaeger and Ted Kaehler, and the Koko
software by Larry Yaeger, calling attention to Koko's role as our advisor
on animal behavior.
Another continuing strand in the Vivarium Program may be thought of as demonstrations
of concept, and Alan talked about the Evans & Sutherland CT6 Kelp Forest
simulation work. He then finished with the Educom tape, which uses the Kelp
Forest footage, noting the new interface organized around projects rather
than applications, the lack of menus entirely, and the use of gestural input.
As usual, Alan got some very enthusiastic responses from the audience.
Mitch Resnick (MIT) - "*Logo: A Children's Environment for Exploring
Self-Organizing Behavior"
Mitch discussed his implementation of an extension to Logo, called *Logo,
that runs on the Connection Machine, and directly supports the modeling
of simple ecologies. He enthusiastically discussed his reasons for wanting
to do this: to help children experiment with and develop an understanding
of self-organizing behaviors, to provide an ability to show children examples
instead of trying to define terms, and by these methods to give children
a deeper understanding of the concepts by the very act of their construction
of these systems. Mitch called this approach to education "constructionism",
and referred to Piaget's theories on building knowledge through what he
called "constructivism" (In terms of one of Alan Kay's favorite
quotes, "To know the world one must construct it."). Mitch referred
to some of the AL work now going on as true "New Wave Science",
blending analysis and synthesis.
Mitch's extensions to Logo support the programming of the motions of thousands
of turtles (instead of just one), the execution of these turtles' commands
in parallel, the dynamic creation and destruction of turtles, the ability
for turtles to sense one another locally, the ability for turtles to sense
and modify the "environment" around them, and for the environment
itself to be computationally active.
With heart and mind very much in concert with the motivations of the Vivarium
program, Mitch noted a number of the benefits that might accrue to the children
who are allowed to experiment with such a system, including the learning
of new concepts (such as diffusion and randomness), developing useful problem-solving
methodologies (mixing analysis and synthesis), coming to grips with the
sociology of science, and developing a new world view (that incorporates
the realities of cooperation and competition, group dynamics, and other
expressions of self-organization). He also noted a benefit that accrues
to the developers of systems such as these, in these kinds of environments,
namely that dealing with and making things more accessible to children forces
you to think about and respond to the core issues of your problem area.
While *Logo's actual language syntax looked a little bit arcane (such things
as a preceding "@" denoting a neighborhood/environment variable,
and preceding quotation mark denoting a turtle variable), his built in distinction
between turtle and neighborhood, with supporting program-block constructs
("to turtle-step ... end" and "to neighborhood-step ... end")
seem like useful, simplifying abstractions that will help the children in
their model-building.
Panel on the Future of Artificial Life with Norm Packard, Rod Brooks, Doyne
Farmer, Alan Kay, Chris Langton and Mark Pauline
NP chaired the panel and kicked it off by asking everyone to comment
on what they perceived to be the future of AL in both the short term and
the long term. He also wondered aloud just what were likely to be the key
issues regarding an artificial organisms' rights.
AK remarked that people in AL research are actually trying to do
something in the same vein as Art, referring to the Greek view of Art as
an imitation of life. He also noted, responding to NP's comment, that it
isn't necessary to worry about legal rights for ALife forms for now... AL
at this stage is still more like a puppet show; they are mirrors that help
us understand life.
* CL took the visionary road, and stated that ALife is only Artificial
in terms of being made by man, or the type of material used. The Life is
real. This conference is a metaphor for AL, moving from physics to chemistry
to simple replicators to simple organisms to complex organisms to self-organizing
AL conferences. Short term he predicts simple extensions of what we have
now. Medium term (100 to 1000 years) humans and ALife will grow in symbiosis.
Long term, he sees a transition to other life forms (evolution does not
stand still).
In response to a comment from the floor, Chris made a rather moving comment
about the beginnings and the intended directions of this field: Noting that
the first AL conference (organized by Chris) was held at Los Alamos National
Laboratory, the birth place of nuclear weapons, which were developed in
an atmosphere of 1) secrecy, 2) without consideration for the consequences
and 3) with a specific (military) intent... by conscious design and deliberate
contrast, the AL1 conference reversed all 3 of these axes to work in an
atmosphere of 1) open accessibility to all, 2) trying to be aware of possible
consequences, and 3) without any single goal, especially no military goal.
RB believes it is impossible to predict the long term future. Short
term we will see infiltration of robots into day to day life the way microprocessors
have done. It'll start with toys, then will come smart doors, then communicating
household items, then small robots will populate the world in symbiosis
with man.
MP says we're lucky that the entire AL field is essentially outside
both state science and mass entertainment. This gives science a chance to
re-integrate with art. He also wonders down stream what we are going to
do to entertain the machines...
DF wonders if there is really going to be a useful theory of emergent
complexity, or whether we will just have to add hack to hack to hack. He
felt there were better demos at AL1, but that there was better theory this
time. AL should be done in the most peaceful way possible. He then commented
on some of the early literary references to AL: genetic engineering in Olaf
Stapledon's Last and First Men (1930), Aldous Huxley's Brave New
World in the '30's, J.D. Bernard's The World of the Flesh and the
Devil (and he praised this particular work highly), and, of course,
Mary Shelley's Frankenstein. He also made the remark that evolution
seems to accelerate constantly, so we'd better get set for Mr. Toad's Wild
Ride. (I wonder if this is true, or if evolution might ultimately yield
to the classic logistic curve after all.)
Artificial Life/Artificial Night: An Evening of Demonstrations and Performance
for the Conferees and General Public
On the evening before the final day's talks, an Artificial Art night was
held that was open to both conference attendees and the local civilians.
Mitch Resnick showed some LEGO-based Braitenberg Vehicle/Creature construction
kits. Slides showing a wonderful series of fine art pieces based on completely
fabricated, but painstakingly detailed creatures from other planets, based
on alternative chemical compounds and processes (referred to as the Sulpher
Creatures) were presented by their designer/ sculptor (some of these suckers
were 6 meters tall) Louis Bec. Doctor Skitzenheimer, aka Peter Oppenheimer,
gave a glimpse into an artificial menagerie, a surreal environment, and
real madness. Rudy Rucker demonstrated some beautiful emergent Zhabotinsky
style reactions in his CA program for the IBM PC. Steve Strassman showed
the Zeltzer/McKenna film trailer for "Grinning Evil Death". Heaven
help us, Jonathan Post read some of his poetry. And Mark Pauline and two
other collaborators from Survival Research Laboratories showed some videos
from their macabre performance art works (these are the folks that design
and build lots of malevolent mechanical automatons and then set them up
each other in the name of Art), and attempted but failed to fire a sonic
cannon they had brought with them. (They did succeed in getting it fixed
before the end of the conference the next day - by cannibalizing an old
dishwasher for the needed solenoid - and ended the conference "with
a bang". This thing shook a cloud of dust out of the ceiling, vibrated
everybody's teeth, and scattered one researcher's pile of notes - at the
extreme opposite end of the hall - to the winds.) A good time was had by
all.
The Poster Sessions
There was one poster paper that was so impressive I just had to include
it here...
* Greg Werner and Michael Dyer (UCLA) - "Evolution of Communication
in Artificial Organisms"
This was simply one of the most clever experimental designs I have seen.
And I actually think that Werner and Dyer have demonstrated the evolution
of at least a simple form of one-way communication between artificial organisms.
Their simulated world consists of a simple grid into which "female"
and "male" organisms of the same species are introduced. The female
organisms are immobile, but can produce a "sound". The male organisms
are mobile, but are blind. The male must find the female in order to procreate.
Both creatures are modeled by simple Neural Nets. The female has the male's
location as one of her inputs (she can "see" him) and sound as
her output. The male has the sounds produced by the female as one of his
inputs (he can "hear" her) and directional movement as his output.
Quoting from their abstract, "Because of the strong selection pressure
in this environment to communicate, a system of communication gradually
evolves in the population such that the sounds made by 'speaking' animals
correspond to actions the 'listening' animals should make in order to find
a mate." The female tells the male where to go.
Greg mentioned that their next plans are aimed at introducing selection
pressures to cause the animals to develop internal models of their environment.
I hope to communicate with him and Michael further on this. (At the risk
of over-emphasizing my current prejudices, frankly, it is nice to see Michael
Dyer not only shed of his Shank/symbolic-AI historical trappings, but making
such a significant contribution to connectionist/AL research.)
The Artificial 4H Awards
Blue-ribbons were awarded to the following artificial contestants:
Best Primordial Soup - Gerald Joyce
Bugs That Learn to Like It - David Ackley
Behaviors That Have the Longest Transients - Kristian Lindgren
Best Urban Planning - James Kalin & (programmer of SimCity)
Most Sophisticated Hardware Bugs - Rod Brooks
Most Sophisticated Software Bugs - Mike Travers
Best Circus & Most Thought Provoking Talk - Mark Pauline
* After all is said and done...
The body of work reviewed here is unique and, to my mind, amazing. In preparing
this extensive review, I've seen connections between various presentations
that were not obvious at the first. It certainly serves as wonderful food
for thought as I commence the construction of my own simulated ecology.
Issues I've been pondering regarding the senses of my organisms, their form,
their neural architecture, their genetic structure, environmental physics,
their ability to affect as well as react to their environment, relations
between innate and learned behaviors, possibilities for communication mechanisms
between organisms (perhaps even between experimenter and organism)... all
have been addressed to some degree in these presentations. Though Geoff
Hinton's poignant list of "theories of mind" through the ages
(with each age believing that at last they had finally sorted it out) teaches
a valuable lesson in perspective, perception, and reality, forgive me if
I take considerable pleasure in at least temporarily embracing a belief
that we live in a time when the nature of mind and life itself may finally
begin to yield up some of their secrets.
An e-mail distribution list for the AL community has been set up. To request
inclusion in this list, send mail to [email protected].
Contributions to the complete mailing list may be sent to [email protected].
[The list was moved to UCLA for a while, but has been dormant for a long
time now. - larryy 4/16/96]
Finally, quoting from Chris Langton's preface to the proceedings of the
first AL conference, "Perhaps ... the most fundamental idea to emerge
at the workshop was the following: Artificial systems which exhibit lifelike
behaviors are worthy of investigation on their own rights, whether or not
we think that the processes that they mimic have played a role in the development
or mechanics of life as we know it to be. Such systems can help us
expand our understanding of life as it could be. By allowing us to
view the life that has evolved here on Earth in the larger context of possible
life, we may begin to derive a truly general theoretical biology capable
of making universal statements about life wherever it may be found and whatever
it may be made of."
|
|
|